

A new study (Powers et al., 2026) just confirmed something that should change the way we talk about diet and health—not just heart health, but brain health, metabolism, and, if you’ve been following this site, endocannabinoid health, too.
When you look at red‑blood‑cell fatty acids, you’re not seeing a niche nutrition marker. You’re seeing a master metabolic biomarker: a snapshot of the fat building your cell walls and feeding your signalling molecules.
Powers et al. (2026) analysed red‑blood‑cell membranes from more than 7,000 Americans aged six and up. Over half had an undesirably low Omega‑3 Index—EPA and DHA together below 4% of membrane fatty acids—and only about 2% reached the >8% range linked to lower cardiac risk. Children and teens did even worse, averaging 3.6% (Powers et al., 2026; Harris et al., 2021).
How the Western Diet Loads the Membrane
Here’s the important point: this is direct measurement of what’s in people’s cell membranes, not what they remember eating.
Other countries show a similar pattern. In a German clinical cohort, average Omega‑3 Index was around 5–6% (Yu et al. ,2026), still below the >8% range associated with less heart risk. By contrast, older healthy Japanese had Omega‑3 Index values close to 10%, with much less arachidonic acid (AA) and a more favourable DHA:AA (Hashimoto et al., 2025).
So Western membranes aren’t just low in omega‑3. They’re built around a very different balance of omega‑6 and omega‑3, with AA playing a much bigger role (Powers et al., 2026; Yu et al., 2026; Hashimoto et al., 2025; Blasbalg et al., 2011).
Arachidonic acid: not just “another fat”
If you look at the Powers box‑and‑whisker plot, AA is the second most abundant fatty acid in American red‑blood‑cell membranes, right behind palmitic acid.
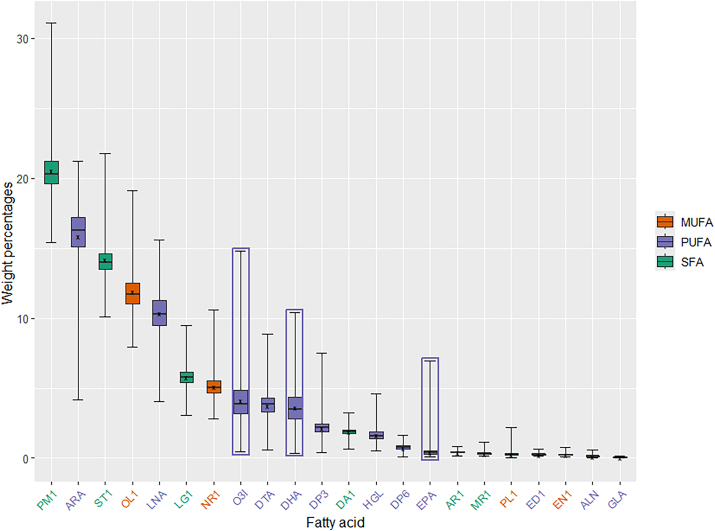
Arachidonic acid is not inert scaffolding. It is the starting material for:
- our two main endocannabinoids, anandamide and 2-AG.
- a whole family of inflammatory molecules: prostaglandins, leukotrienes, thromboxanes (Rouzer & Marnett, 2011; Wang et al., 2021; Tallima and El Ridi, 2017).
Every time a cell needs those signals, it cuts AA out of the membrane. The membrane isn’t the signal; the membrane sets the range of signals that are possible.
The NHANES data tell us what’s sitting in the membrane. They don’t directly tell us how often those signals fire or how many receptors are present. But both of those also depend on diet.
The Western Diet and ECS Tone
When AA is released from the membrane, it reaches a fork in the road.
Down one path, enzymes like COX and LOX turn AA into eicosanoids—molecules linked to inflammation, fever, pain, and immune activation. Down another path, other enzymes turn AA into anandamide and 2‑AG—the endocannabinoids that help control synapses, immunity, pain resolution, and metabolism via CB1 and CB2 receptors (Rouzer & Marnett, 2011; Innes & Calder, 2020).
Same substrate, different outcomes. Which branch dominates depends on the enzymes, the inflammatory state, and how much AA the membrane is holding.
Alvheim et al. showed what happens when you flood this fork with too much substrate (Alvheim et al., 2012). Increasing dietary linoleic acid (LA) raised tissue AA, boosted 2‑AG and anandamide, and led to obesity and inflammation—even on a low‑fat, isocaloric diet. Adding EPA/DHA shrank the AA pool and normalised endocannabinoid tone.
More recently, Liu et al. found that insulin spikes increase the number of CB1 receptors in the liver by slowing their breakdown (which I wrote about in more detail here). A normal‑size insulin pulse can push hepatic CB1 protein up by about 50% in a few hours.
Put these together:
- Poor fat quality (high linoleic acid, low EPA/DHA) loads the membranes with AA and biases signalling toward omega-6-derived endocannabinoids and eicosanoids (Powers et al., 2026; Innes & Calder, 2020; Alvheim et al., 2012).
- Poor carbohydrate quality (frequent high-glycaemic hits) sends repeated insulin spikes that stabilise CB1 receptors and increase receptor numbers on the cell surface (Liu et al., 2026).
In the liver—where CB1 activation promotes fat build‑up and worsens insulin signalling—that’s the last combination you want: more ligand plus more receptor in the wrong organ (Osei-Hyiaman et al., 2008; Liu et al., 2026).
Beyond heart health: brain and pain
Most people hear “Omega‑3 Index” and think heart disease. The same membrane biology applies in the brain and pain pathways.
Consider what we now know about AA in the context of the brain.
AA is one of the main fatty acids in neural tissue and the substrate for both endocannabinoids and neuroinflammatory eicosanoids (Tallima & El Ridi, 2018). A chronically large AA pool plus depleted EPA/DHA tilts brain signalling toward more prostaglandin E2, more microglial activation, and more diversion of tryptophan toward kynurenine instead of serotonin (Wang et al., 2021; Tallima & El Ridi, 2017).
Pain data point in the same direction. Human cohorts show that higher n‑6:n‑3 ratios and higher LA/AA are linked to lower pain thresholds and more pain conditions. A membrane richer in omega‑6 substrates appears more permissive of inflammatory pain signalling (Sanders et al., 2022; Sanders et al., 2023).
Connect that back to NHANES (Powers et al., 2026) —AA as the second most abundant RBC fat, omega‑3 chronically low—and a sobering conclusion emerges:
we are living in a population with lower pain thresholds baked into its membranes.
The prenatal membrane risk gradient
Al-Sinani et al. (2026) measured fatty acids in mothers and their newborns. AA rose from about 13.6% in maternal red‑blood‑cell membranes to 14.9% in fetal cells, while DHA barely moved. LA was lower in fetal blood, meaning the placenta is actively concentrating AA and excluding LA. Ratios like AA/LA and AA/DHA were higher in fetal cells, showing placental AA biomagnification
At the same time, a systematic review by Sarantaki et al. found that maternal low ω‑3:ω‑6 ratios and low DHA, plus cord‑blood AA‑derived oxylipins and lipidomic disturbances, were prospectively linked to more autistic traits and ASD with intellectual disability in the cohorts studied (Sarantaki et al., 2025).
So the picture is:
- Western diets push women’s membranes toward AA‑rich, omega‑3‑poor profiles.
- The placenta then preferentially moves AA into the fetus.
- Perinatal lipidomics now show that this kind of omega‑6‑heavy, DHA‑poor background is a reproducible risk signature for neurodevelopmental problems, including ASD.
RBC fatty acids as a “membrane HbA1c”
If you zoom out, a single RBC fatty‑acid panel is doing several jobs at once:
- It shows the substrate reservoir for endocannabinoids and eCBome mediators (AA → AEA/2‑AG; oleic → OEA; palmitic → PEA; DHA → DHEA).
- It reflects aspects of fat oxidation and peroxisomal load.
- It acts as a proxy for PPAR tone, because many long‑chain fatty acids are direct PPAR ligands (Innes & Calder, 2020).
In that sense, RBC fatty acids behave like an HbA1c for the lipid‑signalling world: a multi‑month average of what your membranes are made of and which signals they can generate.
Mainstream medicine currently treats them as mostly a heart‑risk tool. That’s a missed opportunity.
The protein craze is a distraction
There is a cultural moment happening right now around dietary protein. Protein content is on everything. High-protein versions of traditionally carbohydrate-heavy foods fill supermarket shelves. The message is simple and, from a marketing perspective, effective: eat more protein, build more muscle, live better.
None of that is necessarily wrong. But it is crowding out a conversation we urgently need to have:
the quality of dietary fat matters at a mechanistic, membrane-level, cell signaling level that we are only beginning to properly describe and measure.
You can eat a perfect amount of protein, hit your gym targets every day, and still be living in the 54%—with an undesirable Omega-3 Index and an AA-saturated membrane driving chronic inflammatory signaling from every cell in your body. Protein content tells you nothing about what your cell walls are made of. Saturated-fat content tells you nothing about the AA:EPA balance your immune cells and neurons are drawing from. Caloric density tells you nothing about whether your ECS substrate is biased toward resolution or inflammation.
The Powers study is using the gold-standard measurement—direct RBC membrane fatty-acid quantification—and it is finding a population-wide deviation from anything resembling an optimal lipid profile. That finding deserves to land with the same weight as a study finding that 54% of Americans have undesirable cholesterol levels. It won’t, because we have never agreed on what “optimal” looks like for long chain fatty acids in erythrocyte membranes.
The fat‑1 mouse literature shows what a very different omega‑6/omega‑3 membrane balance can look like: lower AA, higher EPA/DHA, a cooler ECS, and broad shifts in microbiome, metabolomics and disease risk. I unpack that multi‑omic story in more detail in a separate post here.
The politics of “optimal” membranes
What sits between the Powers baseline and the fat‑1 target is not just a technical gap; it is a political and commercial one.
Regulatory reference ranges are not neutral. The moment you draw a line on a lab report and label it “optimal AA:EPA” or “AA above this range increases long‑term cardiometabolic and neurodegenerative risk,” you are no longer talking about abstract biochemistry—you are talking about product categories. A validated reference interval for linoleic‑acid–driven AA loading would, by definition, turn a very large fraction of the current ultra‑processed food supply into something that can be described, in clinical language, as membrane‑distorting.
That is not a conversation Big Food is eager to have. If we agreed, for example, that an AA:EPA ratio above a certain threshold or an O3I below 8% reflects an unhealthy membrane state, then the entire business model built on cheap seed oils and “fortified” high‑margin products starts to look like what it really is: a slow, structural intervention into population lipid signaling. The profit margins on those oils depend on their invisibility at the biomarker level. A world in which every health check includes an AA:EPA ratio and a clear “out of range” flag is not a friendly world for companies whose formulations push that ratio in the wrong direction. That does not require a conspiracy; it only requires inertia in a system where the most profitable ingredients are also the ones most likely to push AA and LA in the wrong direction.
So the missing reference ranges are not just an oversight. They are the absence of a yardstick that would force us to name a lot of current products—and whole categories of “heart‑healthy” snacks—for what they do to membranes over time. The Powers paper gives us the national baseline. The fat‑1 literature gives us a biologically coherent target. The step in between—translating that into official, clinically actionable ranges for membrane fatty‑acid composition—is conspicuously absent, and the longer it stays absent, the more it serves incumbent interests rather than public health.
What you can do in the meantime
The biology points in one direction consistently. Move your membrane toward a lower AA:EPA ratio and higher DHA.
- Reduce linoleic acid intake first — the primary driver of membrane AA accumulation over years of Western eating. That means reducing industrial seed oils in cooking and processed foods; olive oil, butter, and animal fats have far lower linoleic-acid content (Alvheim et al., 2012; Blasbalg et al., 2011).
- Add pre-formed EPA and DHA — oily fish two or more times per week, or a quality marine omega-3 supplement. The ALA conversion route from flax is slow and genetically constrained (Harris et al., 2021).
- Measure your membrane, not just your diet — the Omega-3 Index gives you a real signal. Most people in the Western world score around 4–5%; prospective cohorts suggest above 8% is where meaningful protection begins (Powers et al., 2026; Harris et al., 2021).
- Tame your insulin spikes — shift carbohydrates toward lower-glycaemic sources, add fibre and protein to mixed meals, and avoid constant snacking. The Liu study shows that every big insulin pulse is also a CB1-stabilising pulse in the liver; fewer, smaller spikes mean fewer CB1 receptors sitting primed on the hepatocyte surface (Liu et al., 2026).
The steps I’m summarising here are the same ones the fat‑1 mouse work points to, and they’re consistent with human trial data showing that shifting RBC fatty acids can modulate ECS markers. I discussed that human trial—and why “low plasma LA” is not a licence to drink seed oil—in more detail here.
The conversation about dietary fat quality is long overdue. The NHANES data just gave it a nationally representative foundation.
References
Al-Sinani, M., Johnson, M., Crawford, M., & Al Maqbali, M. (2026). Arachidonic acid is preferentially biomagnified over DHA in fetal erythrocytes: Evidence from 172 paired maternal-cord samples in Oman. Prostaglandins, Leukotrienes and Essential Fatty Acids, 208, 102726. https://doi.org/10.1016/j.plefa.2026.102726
Alvheim AR, Malde MK, Osei-Hyiaman D, et al. Dietary linoleic acid elevates endogenous 2-AG and anandamide and induces obesity. Obesity (Silver Spring). 2012;20(10):1984-1994. doi:10.1038/oby.2012.38
Blasbalg TL, Hibbeln JR, Ramsden CE, Majchrzak SF, Rawlings RR. Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. Am J Clin Nutr. 2011;93(5):950-962. doi:10.3945/ajcn.110.006643
Harris WS, Tintle NL, Imamura F, et al. Blood n-3 fatty acid levels and total and cause-specific mortality from 17 prospective studies. Nat Commun. 2021;12(1):2329. Published 2021 Apr 22. doi:10.1038/s41467-021-22370-2
Hashimoto, M., Matsuzaki, K., Matsuda, C., et al. (2025). Impact of the apolipoprotein E ε4 allele on cognition and omega-3 fatty acid levels in the plasma membrane of red blood cells in healthy elderly Japanese population. Dementia and Geriatric Cognitive Disorders Extra, 15(1), 140–151. https://doi.org/10.1159/000548369
Innes JK, Calder PC. Marine Omega-3 (N-3) Fatty Acids for Cardiovascular Health: An Update for 2020. Int J Mol Sci. 2020;21(4):1362. Published 2020 Feb 18. doi:10.3390/ijms21041362
Liu, S. Y., Ting, C. H., Hwang, G. S., et al. (2026). Insulin increases expression of cannabinoid receptor 1 by suppressing lysosomal degradation via ERK signaling pathway. International Journal of Medical Sciences, 23(6), 2027–2038. https://doi.org/10.7150/ijms.126308
Osei-Hyiaman, D., Liu, J., Zhou, L., Godlewski, G., Harvey-White, J., Jeong, W. I., Bátkai, S., Marsicano, G., Lutz, B., Buettner, C., & Kunos, G. (2008). Hepatic CB1 receptor is required for development of diet-induced steatosis, dyslipidemia, and insulin and leptin resistance in mice. Journal of Clinical Investigation, 118(9), 3160–3169. https://doi.org/10.1172/JCI34827
Powers, C. D., Sternberg, M. R., Mineva, E. M., Clifton, C. C., Leachman, J. R., Wambogo, E., & Pfeiffer, C. M. (2026). Over half of the United States population had an undesirably low Omega-3 Index based on erythrocyte membrane measurements: Results from the cross-sectional NHANES from August 2021 to August 2023. Current Developments in Nutrition, 10(6), 107715. https://doi.org/10.1016/j.cdnut.2026.107715
Rouzer CA, Marnett LJ. Endocannabinoid oxygenation by cyclooxygenases, lipoxygenases, and cytochromes P450: cross-talk between the eicosanoid and endocannabinoid signaling pathways. Chem Rev. 2011;111(10):5899-5921. doi:10.1021/cr2002799
Sanders AE, Weatherspoon ED, Ehrmann BM, et al. Circulating polyunsaturated fatty acids, pressure pain thresholds, and nociplastic pain conditions. Prostaglandins Leukot Essent Fatty Acids. 2022;184:102476. doi:10.1016/j.plefa.2022.102476
Sanders AE, Weatherspoon ED, Ehrmann BM, et al. Circulating Polyunsaturated Fatty Acids and Pain Intensity in Five Chronic Pain Conditions. J Pain. 2023;24(3):478-489. doi:10.1016/j.jpain.2022.10.008
Sarantaki A, Ghanchi A, Vermeulen J, et al. Maternal and cord blood lipidomics as predictors of autism spectrum disorders: A systematic review. Metabol Open. 2025;28:100403. Published 2025 Oct 3. doi:10.1016/j.metop.2025.100403
Tallima H, El Ridi R. Arachidonic acid: Physiological roles and potential health benefits – A review. J Adv Res. 2017;11:33-41. Published 2017 Nov 24. doi:10.1016/j.jare.2017.11.004
Wang, B., Wu, L., Chen, J., Dong, L., Chen, C., Wen, Z., Hu, J., Fleming, I., Wang, D. W., & Cao, Y. (2021). Metabolism pathways of arachidonic acids: Mechanisms and potential therapeutic targets. Signal Transduction and Targeted Therapy, 6, 94. https://doi.org/10.1038/s41392-020-00443-w
Yu, Y., Wang, C., Wang, R., et al. (2026). Age-related alterations in fatty acid metabolism: A clinical study of erythrocyte and plasma profiles in a population from Brandenburg, Germany. Frontiers in Aging, 7, 1725187. https://doi.org/10.3389/fragi.2026.1725187