

There is a mouse that refuses to follow the script. Feed it a high-fat Western diet for sixteen months and it stays leaner, its gut stays tighter, its blood stays cleaner, and its cells age more slowly. Not because of a drug. Because of one change: the balance of fats in its cell membranes.
This mouse is called fat-1. And if you understand why it thrives, you understand something profound about your endocannabinoid system — and about your next meal.
In this article, we use the fat-1 mouse endocannabinoid system model to show how the fats in your cell membranes shape metabolism, inflammation, and long‑term health.
Your ECS Runs on Membrane Fat
The two main endocannabinoids — anandamide and 2-AG — are not stored anywhere. They are made on demand, by clipping molecules out of the cell membrane wall. The raw material is whatever fatty acid happens to be sitting in that wall at the time.
In a modern Western diet, the fatty acid that dominates is linoleic acid — the omega-6 fat in sunflower, corn, and soybean oil. The body converts linoleic acid into arachidonic acid. And from arachidonic acid, it makes both anandamide and 2-AG.
The consequence is direct: the more linoleic acid has accumulated in your membranes over months and years, the larger the pool your ECS draws from — whether you actually need the signal or not.
For example, In 2012, Alvheim and colleagues tested this directly. They raised dietary linoleic acid from 1% to 8% of calories — mirroring the shift in Western diets since the 1960s. That single change elevated tissue arachidonic acid, drove up liver 2-AG and anandamide, and induced obesity, without any change in total caloric intake. The authors concluded that normalising the AA-phospholipid pool by consuming sufficient EPA and DHA was the lever that could reverse it.¹
The membrane is the fuel tank. The diet fills it. The ECS draws from it.
What Happens as the Omega-6/Omega-3 Ratio Shifts
A 2016 study by Su and colleagues makes the dose-response curve visible. They measured how fatty acid composition changes in the liver, brain, and red blood cells across three dietary omega-6/omega-3 ratios: 1:1, 7:1, and 44:1.²
The pattern is the same in all three tissues. As the dietary ratio climbs, arachidonic acid fills the membranes — nearly tripling in the liver between 1:1 and 44:1. Meanwhile, DHA and EPA are physically displaced, competed out of membrane phospholipids by the incoming omega-6.
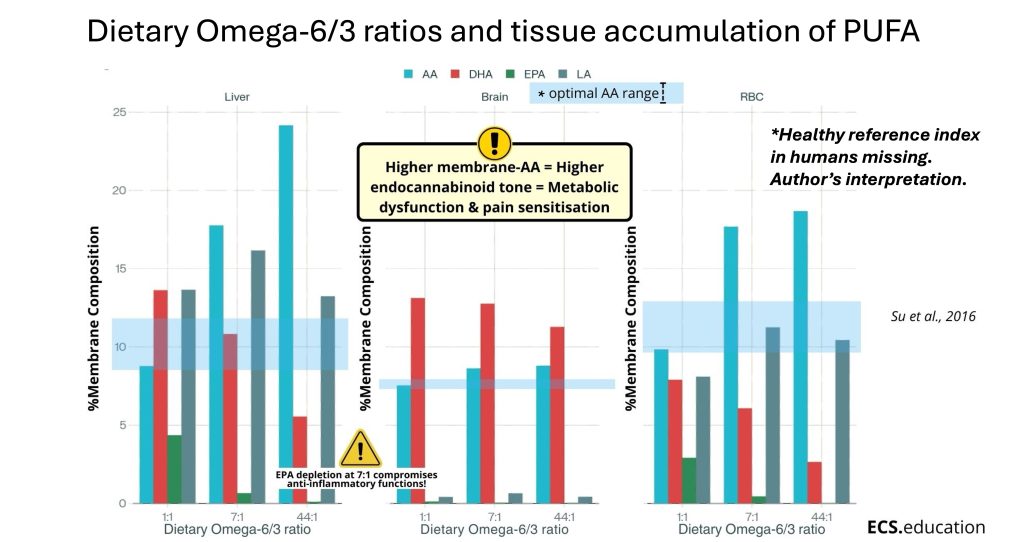
Most striking, the number that should stop you is the one at 7:1. That dietary ratio is still considerably better than the modern Western average of roughly 15–17:1.³ And yet, at 7:1, EPA in the liver is already near zero. That matters because EPA is the membrane substrate for anti-inflammatory and pro-resolving signalling. Lose it, and you do not just have more pro-inflammatory drive — you lose the system’s ability to resolve inflammation at all.
What Su et al. measured is membrane remodelling.
Why this matters for inflammation
What the endocannabinoid data allow us to conclude is what that remodelling means: as arachidonic acid fills the membrane, the ECS has more substrate to draw from — more 2-AG and anandamide on demand, whether the signal is needed or not.
A note on interpretation: the causal link between membrane-AA levels and ECS tone is the author’s synthesis, integrating Su et al.’s membrane data with endocannabinoid biochemistry. No validated human reference range for optimal membrane AA composition currently exists — which is precisely the problem the policy section of this article addresses.
One more thing from this study is worth carrying forward: the red blood cell curves track the liver and brain curves closely. That is the scientific basis for using the Omega-3 Index — EPA and DHA as a share of red blood cell membranes — as a practical window into the tissues that matter most.²
The Fat-1 Mouse Endocannabinoid System in Reverse
The fat-1 mouse carries a gene from a roundworm that does something no mammal can normally do: convert omega-6 fats into omega-3 fats inside its own cells.⁴ Feed it a Western diet and every tissue quietly fills with EPA and DHA while arachidonic acid recedes. In Su et al.’s terms, it lives permanently in the left-hand column.
In 2016, Demizieux and colleagues asked: what does that membrane environment do to the endocannabinoid system? The answer was striking. Fat-1 mice had more than 80% less 2-AG and around 50% less anandamide in their livers than wild-type mice on the same food. Same enzymes. Same calories. Only the membrane composition differed.⁵
In parallel, Kaliannan’s data show that this ECS suppression corresponded to a 50% reduction in liver membrane arachidonic acid — from 6.4% in wild-type to 3.2% in fat-1 mice.7 The relationship is non-linear: a halving of membrane AA substrate produced an 83% collapse in 2-AG, suggesting the ECS amplifies membrane composition changes into proportionally larger shifts in endocannabinoid tone.
This is the core insight of this article: the ECS is not just a receptor system — it is a membrane system. What you build your cells from determines what signals they generate.
What this means for ordinary diets
But the Demizieux study contains a second finding that is, if anything, more important for human application. Alongside the transgenic mice, the researchers fed ordinary wild-type mice a diet enriched with linseed oil — a source of ALA, the plant omega-3 found in flaxseed and walnuts. No EPA. No DHA. Just a better dietary ratio.
Those mice showed the same marked reduction in liver endocannabinoid tone as the fat-1 mice.
The transgenic mouse proves the mechanism. The linseed-fed mouse proves you can get there from a dinner plate.
There is a third finding: some of the protective gene expression changes in the liver persisted even after the mice were switched to a high-fat challenge diet, and were associated with improved glycaemic control. Early omega-3 exposure left a metabolic memory — a partial resistance to later insult encoded in liver gene expression that outlasted the dietary intervention itself. The authors concluded that early n-3 fatty acid interventions “may represent an alternative strategy to drugs for reducing endocannabinoid tone and improving metabolic parameters in the metabolic syndrome.”⁵
Human Evidence: Flaxseed Oil and the Endocannabinoid System
Animal studies prove mechanisms. Randomised controlled trials in humans test whether those mechanisms operate in the real world. In 2020, Saleh-Ghadimi and colleagues did exactly that.⁶
They enrolled 44 patients with coronary artery disease — a population with chronically elevated ECS tone and systemic inflammation — and gave half of them flaxseed oil providing 2.5 g of ALA per day for ten weeks. The other half received placebo.
The membrane changed first. Arachidonic acid in red blood cell membranes fell by more than 4 percentage points within the intervention group. Linoleic acid fell too. The omega-6 load in the cell wall measurably declined.
Then the ECS followed. Circulating anandamide fell significantly in the flaxseed group compared to placebo. And CB2 — the receptor subtype associated with anti-inflammatory, peripheral signalling rather than the metabolically overactive CB1 profile — increased its expression by 30%.
In ten weeks. With flaxseed oil. In people with heart disease.
This closes the translational chain: dietary ALA shifts the membrane, a shifted membrane changes ECS tone, and a changed ECS tone moves the body toward a less inflammatory, less metabolically stressed state. The fat-1 mouse showed what the destination looks like. The Demizieux mice showed a dietary path toward it. Saleh-Ghadimi showed it works in humans.
When the Whole System Shifts
A 2019 Harvard-led study asked what happens when you measure not just one organ, but the entire organism at once.⁷
Four mouse lines — spanning from genetically high omega-6 to fat-1-like omega-3 enrichment — all ate the same Western diet for sixteen months. The researchers measured gut bacteria, hundreds of metabolites in stool and blood, immune markers across multiple tissues, gut barrier integrity, and disease development simultaneously.
The high omega-6 mice arrived at a recognisable place. Their gut walls became leaky. Bacterial toxin entered the bloodstream. Immune alarm receptors fired in the gut, liver, and fat tissue. And across fecal and blood metabolites, the team found marker after marker already validated in human studies as a signal of chronic disease: molecules linked to liver disease, insulin resistance, type 2 diabetes, atherosclerosis, and cancer risk.
Two very different biological worlds
In contrast, the fat-1-like mice told the opposite story. A tighter gut wall. A protective microbial composition. Anti-inflammatory metabolites, including indolepropionate — a compound associated with reduced type 2 diabetes risk and better gut barrier function in humans. Dramatically less disease over sixteen months.
When the researchers used network mathematics to integrate all three layers of data — microbiome, metabolites, disease outcomes — the two groups sat in entirely different biological worlds. The authors put it directly: excess omega-6 in tissues is harmful, and a low tissue omega-6/omega-3 ratio reduces chronic disease risk, demonstrated simultaneously across the microbiome, the metabolome, and clinical outcomes.⁷
The membrane shift the fat-1 mouse embodies is not a cardiovascular intervention. It is a whole-organism intervention.
The Brain, Ageing, and the ECS
The same membrane advantage reaches the brain. A 2025 study subjected both wild-type and fat-1 mice to chronic sleep deprivation. Both groups deteriorated — anxiety, memory loss, circadian disruption, brain inflammation, and early Alzheimer-like amyloid changes. The fat-1 mice deteriorated significantly less across every measure.⁸ Same stressor. Different membrane background. Less damage.
In addition, a 2024 study looked further — at telomeres, the protective caps on chromosomes that shorten a little with every cell division and faster still under chronic stress. Fat-1 mice showed slower telomere shortening with age than their wild-type counterparts.⁹ The authors described it as the first demonstration, in an animal model free of dietary confounders, that elevated tissue omega-3 status can slow the biological clock at the chromosomal level.
The membrane environment that reduced liver ECS tone by over 80%, reorganised the whole-body disease-risk pattern of blood and stool molecules, and buffered the brain against sleep deprivation also appears to slow ageing itself.
An Uncomfortable Policy Gap
Here is where the science demands honesty.
There are no validated reference ranges for tissue omega-6/omega-3 ratio in humans. We have converging evidence — transgenic models, datasets across genes, microbes, and metabolites, human RCT data — showing that the entire disease-risk architecture of an organism reorganises around this single variable. And we have no clinical target for it.
What we have instead are dietary guidelines recommending 5–10% of daily calories from linoleic acid¹⁰ — guidance formulated primarily around cardiovascular cholesterol endpoints decades ago, with no reference to tissue membrane composition, ECS tone, or the metabolomic disease signatures Kaliannan’s group mapped in detail.
A population-wide membrane experiment
Meanwhile, adipose tissue linoleic acid levels in Americans rose 136% between 1959 and 2008, tracking the rise of industrial seed oils.¹¹ That is a population-wide membrane experiment that nobody formally approved and nobody is formally monitoring. In human cohort data, a higher plasma omega-6/omega-3 ratio is associated with significantly greater risk of dying from any cause, from cancer, and from cardiovascular disease.¹² A 2025 global survey of over 500,000 dried blood spot samples found that the vast majority of the world’s population has insufficient omega-3 status.¹³
Taken together, the biological case is made. The human epidemiology is consistent. The randomised trial data confirms the mechanism. What is missing is the political will to act on it.
We call on policymakers, guideline bodies, and research funders to treat tissue omega-6/omega-3 ratio as a public health priority: to fund long-term human studies with tissue composition as a primary endpoint, to establish reference ranges, and to revisit dietary fat guidance in light of sixty years of membrane biology — not only sixty years of cholesterol science.
What You Can Do Now
Practically speaking, you cannot install the fat-1 gene. But you can move your membranes in the same direction, and the evidence is now clear enough to act on deliberately.
- Reduce the omega-6 load first. This is the highest-leverage change. The Saleh-Ghadimi trial achieved measurable membrane remodelling and ECS attenuation within ten weeks, and the omega-6 displacement from membranes did much of the work. Minimising seed oils in cooking and processed food is where to start.
- Add plant omega-3 sources daily. Flaxseed, chia, and walnuts provide ALA, and both the Demizieux and Saleh-Ghadimi data show ALA alone can shift membrane composition and reduce ECS tone — though how far depends on your FADS2 genetics. If plant sources don’t move your Omega-3 Index, preformed EPA and DHA from oily fish or marine supplements bypass the conversion step entirely.
- Consider preformed EPA/DHA for deeper remodelling. For anyone with cardiovascular disease, metabolic syndrome, or a known poor ALA conversion rate, preformed omega-3 accelerates membrane remodelling and directly replenishes the EPA that disappears from membranes at high dietary ratios.
- Measure your status. The Omega-3 Index gives you a real window into your membrane composition via a simple blood test. Most people in Western countries score around 4–5%. The research consistently points to above 8% as the range where meaningful protection begins.¹⁴
- Move and sleep. Aerobic exercise delivers the kind of ECS activation the system was designed for: transient, rhythmic, resolvable. Chronic sleep deprivation dysregulates the daily ECS rhythm and amplifies the neuroinflammatory damage that a high omega-6 membrane background makes worse.⁸
The fat-1 mouse is not an omega-3 story. It is the closest model we have to a biological system running on the substrate it was designed for — from membrane to chromosome, from gut to brain.
The ECS is a demand-driven, membrane-derived network. What you build your membranes from determines what signals that network generates, how loud they are, and how well they resolve.
Flaxseed oil in a glass of milk. Oily fish twice a week. Less seed oil in the pan. The biology is sophisticated. The path there is not.
References
- Alvheim AR, Malde MK, Osei-Hyiaman D, et al. Dietary linoleic acid elevates endogenous 2-AG and anandamide and induces obesity. Obesity (Silver Spring). 2012;20(10):1984–1994. doi:10.1038/oby.2012.38
- Su H, Zhou D, Pan YX, Wang X, Nakamura MT. Compensatory induction of Fads1 gene expression in heterozygous Fads2-null mice and by diet with a high n-6/n-3 PUFA ratio. J Lipid Res. 2016;57(11):1995–2004. doi:10.1194/jlr.M064956
- Simopoulos AP. An increase in the omega-6/omega-3 fatty acid ratio increases the risk for obesity. Nutrients. 2016;8(3):128. doi:10.3390/nu8030128
- Kang JX. Fat-1 transgenic mice: a new model for omega-3 research. Prostaglandins Leukot Essent Fatty Acids. 2007;77(5–6):263–267. doi:10.1016/j.plefa.2007.10.010
- Demizieux L, Piscitelli F, Troy-Fioramonti S, et al. Early low-fat diet enriched with linolenic acid reduces liver endocannabinoid tone and improves late resistance to diet-induced obesity. Diabetes. 2016;65(7):1824–1837. doi:10.2337/db15-1279
- Saleh-Ghadimi S, Alizadeh M, Jafari-Vayghan H, et al. Effect of flaxseed oil supplementation on the erythrocyte membrane fatty acid composition and endocannabinoid system modulation in patients with coronary artery disease: a double-blind randomized controlled trial. Genes Nutr. 2020;15(1):9. doi:10.1186/s12263-020-00665-1
- Kaliannan K, Li XY, Wang B, et al. Multi-omic analysis in transgenic mice implicates omega-6/omega-3 fatty acid imbalance as a risk factor for chronic disease. Commun Biol. 2019;2:276. doi:10.1038/s42003-019-0521-4
- Niazi ZR [first author TBC] et al. Chronic sleep deprivation induces depression- and Alzheimer’s disease-like changes in adult and ageing wild-type and Fat-1 transgenic mice. Prostaglandins Leukot Essent Fatty Acids. 2025;207:102699. doi:10.1016/j.plefa.2025.102699
- Ogłuszka M, et al. Elevated tissue status of omega-3 fatty acids protects against age-related telomere attrition in fat-1 transgenic mice. Clin Nutr. 2024;43(6):1488–1494. doi:10.1016/j.clnu.2024.05.001
- US Department of Agriculture; US Department of Health and Human Services. Dietary Guidelines for Americans, 2020–2025. 9th ed. 2020.
- Blasbalg TL, Hibbeln JR, Ramsden CE, Majchrzak SF, Rawlings RR. Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. Am J Clin Nutr. 2011;93(5):950–962. doi:10.3945/ajcn.110.006643
- Zhu S, et al. Higher ratio of plasma omega-6/omega-3 fatty acids is associated with greater risk of all-cause, cancer, and cardiovascular mortality. EClinicalMedicine. 2024. doi:10.1016/j.eclinm.2024.102614
- Torrissen M, Gisslevik E, Gundersen TE, et al. Global variations in omega-3 fatty acid status and omega-6:omega-3 ratios: insights from > 500,000 whole-blood dried blood spot samples. Lipids Health Dis. 2025;24(1):260. doi:10.1186/s12944-025-02676-6
- Harris WS, Tintle NL, Imamura F, et al. Blood n-3 fatty acid levels and total and cause-specific mortality from 17 prospective studies. Nat Commun. 2021;12:2329. doi:10.1038/s41467-021-22370-2
Stefan Broselid is a researcher and educator in systems biology and endocannabinoid science at ECS.education
Graphic: Data from Su et al., 2016 | ECS interpretation © ECS.education